ADIPOSE TISSUE
Ann L. Albright and Judith
S. Stern
Department of Nutrition and Internal Medicine
University of California at Davis
Davis, CA
USA
Morphology and
Development of Adipose Tissue
Adipose-Tissue
Metabolism
Adipose Tissue
Distribution
Definition and
Causes of Obesity
Further
Reading
Albright, A.L. and Stern, J.S. (1998). Adipose tissue. In:
Encyclopedia of Sports Medicine and Science, T.D.Fahey (Editor).
Internet Society for Sport Science: http://sportsci.org. 30 May 1998.
Adipose tissue is specialized connective tissue that functions as
the major storage site for fat in the form of triglycerides. Adipose
tissue is found in mammals in two different forms: white adipose
tissue and brown adipose tissue. The presence, amount, and
distribution of each varies depending upon the species. Most adipose
tissue is white, the focus of this review.
White adipose tissue serves three functions: heat insulation,
mechanical cushion, and most importantly, a source of energy.
Subcutaneous adipose tissue, found directly below the skin, is an
especially important heat insulator in the body, because it conducts
heat only one third as readily as other tissues. The degree of
insulation is dependent upon the thickness of this fat layer. For
example, a person with a 2-mm layer of subcutaneous fat will feel as
comfortable at 15°C as a person with a 1-mm layer at 16°C.
Adipose tissue also surrounds internal organs and provides some
protection for these organs from jarring.
As the major form of energy storage, fat provides a buffer for
energy imbalances when energy intake is not equal to energy output.
It is an efficient way to store excess energy, because it is stored
with very little water. Consequently, more energy can be derived per
gram of fat (9 kcal.gm-1) than per
gram of carbohydrate (4 kcal.gm-1)
or protein (4 kcal.gm-1). In
addition, if terrestrial animals stored their excess energy as
carbohydrate, the increased mass would interfere with mobility. The
average woman with 20% body fat has about one month of energy stored
as fat.
There are some constraints on the use of fat as fuel. Most animals
cannot convert lipid into carbohydrate. Tissues that function
predominantly anaerobically (e.g.. erythrocytes) must rely on
carbohydrate for energy and need to have an ample supply available.
Additionally, under normal conditions the brain is dependent upon
glucose for energy and does not use fatty acids. In unusual metabolic
circumstances, the brain can use ketone bodies (a by-product of
incomplete fat metabolism) when they are present in sufficiently high
quantities. Finally, a typical diet contains a high proportion of
carbohydrate, and the transport of insoluble lipids through blood
requires a specific mechanism, so it may be "easier" metabolically
for tissues to use glucose under typical dietary conditions.
Brown adipose tissue, which derives its color from rich
vascularization and densely packed mitochondria, is found in various
locations, depending upon the species and/or age of the animal. In
the rat, brown adipose tissue is found primarily in the interscapular
region and the axillae, minor amounts are found near the thymus and
in the dorsal midline region of the thorax and abdomen. During
maturation, in non-hibernating animals, brown adipose tissue is
metabolically less active, although cold exposure can activate it. In
hibernating animals and neonates, brown adipose tissue is important
for regulating body temperature via non-shivering thermogenesis.
Instead of serving as a substrate, the lipid in brown adipose
tissue releases energy directly as heat and is, therefore used in
heat production for non-shivering thermogenesis and for utilization
of excess caloric intake via diet-induced-thermogenesis. The
mechanism of heat generation is related to the metabolism of the
mitochondria. Mitochondria from brown adipose tissue have a specific
carrier called uncoupling protein that transfers protons from outside
to inside without subsequent production of ATP.
Morphology and Development of
Adipose Tissue
In adult mammals, the major bulk of adipose tissue is a loose
association of lipid-filled cells called adipocytes, which are held
in a framework of collagen fibers. In addition to adipocytes, adipose
tissue contains stromal-vascular cells including fibroblastic
connective tissue cells, leukocytes, macrophages, and pre-adipocytes
(not yet filled with lipid), which contribute to structural
integrity.
|
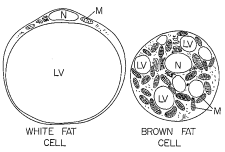
Figure 1. White fat cell and brown fat cell.
Note the single large lipid vacuole in the white fat cell
and the numerous smaller lipid vacuoles in the brown fat
cell. LV: lipid vacuole; M: mitochondria; N: nucleus.
Artwork courtesy of Dr.
John Horwitz, U.C. Davis.
|
|
|
The lipid droplets in adipose tissue can be unilocular and/or
multilocular. Unilocular cells contain a single large lipid droplet
which pushes the cell nucleus against the plasma membrane, giving the
cell a signet-ring shape (Figure 1). Unilocular cells, characteristic
of white adipose tissue, range in size from 25 to 200 microns.
Mitochondria are found predominately in the thicker portion of the
cytoplasmic rim near the nucleus. The large lipid droplet does not
appear to contain any intracellular organelles. Multilocular cells,
typically seen in brown adipose tissue, contain many smaller lipid
droplets. A cell in brown adipose tissue may reach a diameter of 60
microns and the lipid droplet within the cell may reach 25 microns in
diameter. As mentioned above, the brown color of this tissue is
derived from the cells' rich vascularization and densely packed
mitochondria. These mitochondria vary in size and may be round, oval,
or filamentous in shape.
Approximately 60 to 85% of the weight of white adipose tissue is
lipid, with 90-99% being triglyceride. Small amounts of free fatty
acids, diglyceride, cholesterol, phospholipid and minute quantities
of cholesterol ester and monoglyceride are also present. In this
lipid mixture, six fatty acids make up approximately 90% of the
total, and these are myristic, plamitic, palmitoleic, stearic, oleic,
and linoleic. Varying the composition of your diet can vary the fatty
acid profile in adipose tissue. The remaining weight of white adipose
tissue is composed of water (5 to 30%) and protein (2 to 3%).
White adipose tissue is not as richly vascularized as brown
adipose tissue, but each adipocyte in white adipose tissue is in
contact with at least one capillary. This blood supply provides
sufficient support for the active metabolism, which occurs in the
thin rim of cytoplasm surrounding the lipid droplet. Blood flow to
adipose tissue varies depending upon body weight and nutritional
state, with blood flow increasing during fasting.
Adipocytes are considered to originate from fibroblast-like
precursor cells that differentiate into adipocytes under the
appropriate stimulatory conditions (described below). The precursor
cells do not possess any morphological or enzymatic marker that can
be used to determine whether they will become adipocytes. The
criteria used to identify adipocytes depends upon lipid accumulation
within the cell after proliferation has stopped, making early
identification of adipocytes difficult.
The size of adipose tissue mass is a function of both adipocyte
number and size. An increase in adipose tissue mass can occur by
hyperplastic growth, which is an increase in the number of
adipocytes. This increase in number occurs primarily by mitotic
activity in precursor cells. Adipose tissue mass can also increase by
hypertrophic growth, which is an increase in the size of adipocytes.
This increase in size occurs primarily by lipid accumulation within
the cell. Growth of this tissue in the rat occurs in well-defined
stages. From birth to 4 weeks of age, adipose tissue growth is
hyperplastic. Overfeeding a rat during this period can lead to
permanent increases in body weight and fat cell number. From 4 to 14
weeks of age both adipocyte hypertrophy and hyperplasia occur.
Following 14 weeks of age, adipose tissue growth occurs predominantly
by adipocyte hypertrophy.
The developmental sequence of adipose tissue in humans is less
well defined. In contrast to most neonates, the human neonate is born
relatively fat. Two periods of hyperplastic growth are probably
during the third trimester of pregnancy and just prior to and during
puberty.
Contrary to earlier belief, hyperplastic growth can also occur in
adulthood (in both humans and rats). When adipocytes fill with lipid
and get to a critical size, precursor cells are stimulated to
differentiate, and an increase in adipocyte number results. This
critical size probably does not occur with moderate overfeeding
unless the overfeeding is of long duration. In addition, there are
probably individual differences in the size that will result in new
adipocyte formation. Once new adipocytes are formed, they remain
throughout life and only a reduction in size of the cell is possible.
This increased number of adipocytes has far-reaching consequences for
the treatment and prevention of obesity.
Adipose-Tissue
Metabolism
Lipogenesis
Lipogenesis is the deposition of fat. This process occurs in
adipose tissue and in the liver at cytoplasmic and mitochondrial
sites (Figure 2).
|
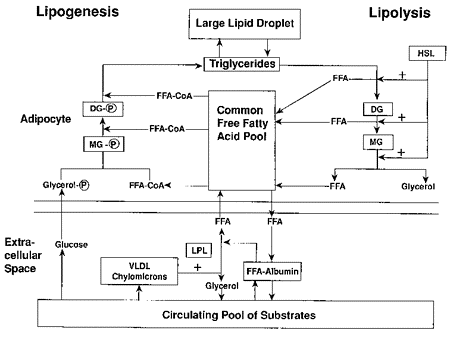
Figure 2:
Diagrammatic
representation of triglyceride storage (lipogenesis) and
breakdown (lipolysis) in adipocytes. FFA: free fatty acid;
FFA-CoA: thioester; HSL: hormone sensitive lipase; LPL:
lipoprotein lipase; DG: diglyceride; MG: monoglyceride;
DG-P: phosphatidic acid; MG-P: lysophosphatidic acid; VLDL:
very low density lipoproteins.
Adapted from Leibel,
Berry, and Hirsch, 1983.
|
|
|
Energy ingested as fat beyond that needed for current energy
demands is stored in adipose tissue. In addition, carbohydrate and
protein consumed in the diet can be converted to fat. Energy ingested
as carbohydrate can be stored as glycogen in the liver and muscle.
Carbohydrate can also be converted to triglycerides primarily in the
liver and transferred to adipose tissue for storage. Amino acids from
ingested proteins are used for new protein synthesis or they can be
converted to carbohydrate and fat.
Fatty acids, in the form of triglycerides or free fatty acids
bound to albumin, are ingested in the diet or synthesized by the
liver (described above). Very little synthesis of free fatty acids
occurs in the adipocytes. Triglycerides are the most significant
source of fatty acids, because this is the form in which dietary
lipids are assembled by the gut and liver. Triglycerides made up of
long chain fatty acids, in the form of chylomicrons (from intestinal
absorption) or lipoproteins (from hepatic synthesis), are hydrolyzed
to glycerol and free fatty acids by an enzyme called lipoprotein
lipase (LPL). Lipoprotein lipase is synthesized in adipocytes and
secreted into adjacent endothelial cells. Chylomicrons and
lipoproteins (very low density lipoproteins) contain C-ll apoprotein,
which activates LPL. Free fatty acids are taken up by adipocytes in a
concentration-dependent manner by a transmembrane transport protein.
Once inside the adipocyte, fatty acids enter a common pool made up of
both incoming and outgoing fatty acids. Fatty acids that are stored
in the adipose tissue must first combine with coenzyme A to form a
thioester and then they are re-esterified in a stepwise manner to
triglycerides. Glucose is the primary source of glycerol for this
re-esterification process. Only a small amount of glycerol released,
when triglycerides are hydrolyzed by LPL, can be reused by adipocytes
to form alpha glycerol phosphate to be used for trigyceride assembly.
Most glycerol is returned to the circulation.
Insulin, a hormone secreted by the beta cells of the pancreas,
plays a predominant role in the lipogenic process. The net effect of
insulin is to enhance storage and block mobilization and oxidation of
fatty acids. Insulin exerts its effect by stimulating LPL formation,
so that circulating triglycerides are hydrolyzed and free fatty acids
can enter the adipocyte. Insulin is also required for the transport
of glucose, which is needed for re-esterification of the
triglycerides once inside the adipocyte. Finally, the conversion of
glucose to fatty acids is accomplished by insulin's activation of
several enzymes.
Lipolysis
Lipolysis is the chemical decomposition and release of fat from
adipose tissue. This process predominates over lipogenesis when
additional energy is required (Figure 2). The triglycerides within
the adipocyte are acted upon by a multi-enzyme complex called hormone
sensitive lipase (HSL), which hydrolyzes the triglyceride into free
fatty acids and glycerol. These lipases act consecutively on
triglycerides, diglycerides, and monoglycerides. Triglyceride lipase
regulates the rate of lipolysis, because its activity is low.
Once triglycerides are hydrolyzed to fatty acids and glycerol,
fatty acids enter the common free fatty acid pool where they may be
re-esterified, undergo beta-oxidation (metabolic degradation), or be
released into the circulation as substrates for skeletal muscle,
cardiac muscle, and liver. If the fatty acids are to undergo
beta-oxidation for ATP production, fatty acids move from the
adipocytes into the blood and are carried to the tissues that can use
them as an energy source. Long-chain fatty acids enter the cells of
these tissues by passive diffusion, and their rate of uptake is
proportional to their difference in concentration inside and outside
of the cell. Once inside the cells, beta-oxidation begins with
"activation", that is, the formation of thioesters with coenzyme A.
This activation step converts the fatty acids to a form that is more
amenable to the successive biochemical changes that ultimately result
in ATP formation.
Insulin reduces mobilization of fatty acids from adipose tissue by
inhibiting triglyceride lipase. The mechanism of this inhibition may
be through a decrease in cyclic AMP which in turn results in an
inhibition of cyclic-AMP-dependent protein kinase. This suppression
of lipolysis lowers the rate of fatty acid delivery to the liver and
to peripheral tissues. The consequence of fewer fatty acids to the
liver is a reduction in the formation of ketoacids (e.g., ketones).
Insulin also stimulates the use of ketoacids by peripheral tissues,
preventing an accumulation of these acids in the blood.
Adipose Tissue
Distribution
|
Figure 3: Representation of "pear" or lower
body fat distribution and "apple" or upper body fat
distribution.
Artwork courtesy of
Wendy M. Garcia, Davis, CA.
|
|
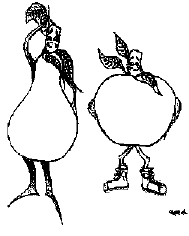
|
Even the most casual observer will note that everyone does
not carry their adipose tissue in the same anatomical locations.
Adipose tissue that is located predominantly on the upper body has
been termed android, male, central, upper-body segment, or "apple"
distribution (Figure 3).
This distribution pattern is found more frequently in men;
consequently the terms android and male. When adipose tissue
accumulates predominantly in the lower body, the terms gynoid,
female, lower-body segment , or "pear" distribution are used (Figure
3). As the terms gynoid and female imply, this fat pattern is found
more frequently in women than in men.
What factors determine fat distribution? A primary factor is
genetic background, which can often be seen by looking at the
similarity in fat distribution within same-sex family members. As
mentioned earlier, gender is also known to affect body fat location.
Women usually show greater lower body fat distribution and men show
more upper body fat distribution. As an individual ages body fat
distribution may change. For example, after the menopause a change
towards upper body fat distribution is observed. This change may be
due to a relative decrease in LPL activity in the lower body region.
Finally, extreme weight gain and weight cycling ("yo-yo dieting") may
increase upper body fat.
Upper body fat distribution is correlated with the development of
various health problems, including cardiovascular disease,
hypertension, and non-insulin dependent diabetes. Excess visceral fat
rather than excess upper body subcutaneous fat is associated with
increased risk. A quick test to see if you are at risk for
complications associated with upper body fat distribution is to
calculate your waist-to-hip-ratio by measuring the circumference of
your waist and dividing it by the circumference of your hips. Women
are at risk if the ratio exceeds 0.85; for men the ratio is 0.95.
Definition and Causes of
Obesity
Obesity is not a single disorder. A variety of methods and
criteria are used to diagnose the presence of obesity. It is the
quantity of adipose tissue and not just total body weight that
defines obesity. Several factors contribute to the development of
obesity: genetics, environment, physiology, psychology, and
undetermined.
Work from experimental animals shows clear genetic anomalies as
one basic cause of obesity. Genes for obesity can be recessive,
dominant, or polygenic. Recessive alleles that influence the
development of obesity have been identified in various rodents.
In the human population, obesity is a feature of several recessive
syndromes, including Bardet-Biedl, Lawrence-Moon, and
Prader-Labhart-Willi, as well as, triglyceride storage diseases.
These syndromes are relatively rare. A single genetic anomaly in the
absence of other concurrent genetic syndromes has yet to be
identified in humans. It is possible that alleles at particular loci
exert a small effect that is not easily detectable by themselves. If
these alleles exist at a large number of loci, collectively they may
exert a substantial impact on obesity. Genes clearly influence body
size and body fat distribution, and are likely to be involved in
modulating human obesity.
In humans, it is difficult to separate genetic and environmental
factors and assign the relative contribution of each to the
development of obesity. Several familial, twin, and adoption studies
have provided a means to study the contributions of heredity and
environment. These studies provide strong suggestive data for a
genetic component to obesity, as well as modulation of genes by
environmental factors.
Environmental influences on obesity include food intake and degree
of physical activity. Overeating relative to energy needs is a
component of obesity development. However, from studies with
experimental animals it is clear that many animals that develop
obesity are energetically more efficient than lean animals; that is,
for a given amount of food, the obesity-prone animals deposit more
fat than do the obesity-resistant animals. In fact, when genetically
obese rodents are not allowed to overeat from birth, they are still
considerably fatter than their lean brothers and sisters.
The study of food intake in humans is confounded by the fact that
some obese individuals under-report the amount of food they eat. The
composition of the diet can also influence obesity. Overweight
individuals have been shown to consume more high-fat foods than
normal-weight individuals.
The amount of energy that an individual expends will also
influence the development of obesity. Increased level of activity is
associated with leanness. Other than the energy deficit that occurs
with exercise, metabolic changes with training also impact fat
utilization and storage. Trained individuals have an increased
lipolytic capacity compared to sedentary persons. Both basal and
catecholamine-stimulated lipolysis are higher in fat cells from
trained individuals. In rats that are obesity-prone, prior to the
onset of obesity, catecholamine-stimulated lipolysis is lower in
their fat cells than in fat cells from obesity-resistant rats. An
important benefit of exercise during weight loss is that it results
in preservation of fat-free mass and allows fat loss that is
equivalent to or greater than weight loss.
Several physiological factors may be involved in the development
of obesity. These include altered adipose tissue metabolism, hormonal
changes, and changes at sites in the brain that control satiety,
particularly in the hypothalamus. There may be some kind of abnormal
signal that affects adipose tissue metabolism and alters fuel
partitioning, directing increased storage in adipose tissue instead
of use in muscle. This abnormal signal has not been identified.
However, lipoprotein lipase has been found to be increased in
obesity, and this enzyme can increase the storage of triglycerides in
adipose tissue. Hormonal changes that may affect obesity include
hyperinsulinemia and changes in pituitary or adrenal function.
Hormonal changes during pregnancy promote fat deposition. If weight
gain is excessive, it can result in lifelong obesity.
The role of psychological variables in the development of obesity
is difficult to define. There is no unifying theme. Some obese
individuals have eating disorders. Other obese individuals may eat in
response to stress or depression. Still other obese individuals may
eat large meals or may eat rapidly. It has also been suggested that
some obese individuals may be restrained eaters, and when there is a
break in their usual routine, they respond by excessive eating.
Although a number of mechanisms have been identified for the
development of obesities, the exact picture is far from clear. For
many individuals, there may be an underlying biological defect. When
this individual comes into contact with a plentiful, good-tasting,
high-fat food in an environment that promotes inactivity, obesity
results. Also important is the fact that while many people may lose
weight, there is little success in keeping the weight off. The
reasons for this recidivism and possible solutions are the focus of
current research.
Further Reading
1. Bouchard C., A. Tremblay, A.
Nadeau, J. Dussault, J.-P. Despres, G. Theriault, P.J. Lupien, O.
Serresse, M.R. Boulay, and G. Fournier. Long-term exercise training
with constant energy intake. 1: Effect on body composition and
selected metabolic variables. Int. J. Obesity 14:57-73, 1990.
2. Greenwood, M.R.C. and P.R.
Johnson. Genetic differences in adipose tissue metabolism and
regulation. Ann. N.Y. Acad. Sci. 676:253-269, 1993.
3. Johnson, P.R. and M.R.C.
Greenwood. The adipose tissue. In: "Cell and Tissue Biology: A
Textbook of Histology." (ed. by L. Weiss), 6th edition, Urban and
Schwarzenberg, Baltimore, MD. pp. 191-209, 1988.
4. Johnson, P.R., J.S. Stern, M.R.C.
Greenwood, L.M. Zucker, and J. Hirsch. Effect of early nutrition on
adipose cellularity and pancreatic insulin release in the Zucker rat.
J. Nutrition. 103:738-743, 1973.
5. Knittle, J.L. and J. Hirsch.
Effect of early nutrition on the development of rat epididymal pads:
cellularity and metabolism. J. Clin. Invest. 47:2091-2098,
1968.
6. St. Jeor, S.T. The role of weight
management in the health of women. J. Amer. Dietetic Assoc.
93:1007-1012, 1993.
7. Stunkard, A.J., J.R. Harris, N.L.
Pederson, and G.E. McClearn. The body-mass index of twins who have
been reared apart. N. Engl. J. Med. 322:1483-1487, 1990.
8. Wadden T.A., and T.B. Van Itallie,
Treatment of the seriously obese patient. The Guilford Press, New
York, 524 pp, 1992.
newseditor=AT=sportsci.org · webmaster=AT=sportsci.org · Encyc
Home · Sportsci Home
· ©1998
Edited and webmastered by Will Hopkins
Last updated 10 July 1998